


Cánh cổng hai nêm và dòng năng lượng vô hình nuôi sống thực vật
1. Nitơ – nền móng của sự sống xanh
Mọi cánh rừng, mọi cánh đồng lúa, mọi tán cây vươn lên bầu trời đều dựa trên một nguyên tố: Nitơ. Không có Nitơ, không có amino acid. Không có amino acid, không có protein. Không có protein, không có enzyme. Không có enzyme, không có sự sống.
Trong khí quyển, Nitơ tồn tại dồi dào dưới dạng N₂. Nhưng phần lớn thực vật không thể sử dụng trực tiếp dạng này. Thứ mà rễ cây có thể hấp thu là các dạng hòa tan trong đất, chủ yếu:
-
NO₃⁻ (nitrate)
-
NH₄⁺ (ammonium)
Ở đất thoáng khí, nitrate chiếm ưu thế. Vì vậy, cơ chế hấp thu NO₃⁻ trở thành một trong những hệ thống sinh lý quan trọng nhất của cây.
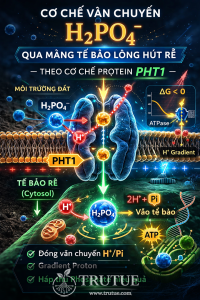
Nhưng hấp thu nitrate không đơn giản như hút nước qua ống. Đó là một bài toán năng lượng, điện tích và cấu trúc protein.
Ở cấp độ phân tử, một protein đóng vai trò trung tâm trong quá trình này là
NRT1.1.
2. Vấn đề nhiệt động học: Vì sao không thể khuếch tán đơn thuần?
Hãy hình dung bề mặt của một tế bào lông hút rễ.
-
Bên ngoài: dung dịch đất.
-
Bên trong: tế bào chất.
-
Giữa hai bên: màng sinh chất.
Điện thế màng của tế bào thực vật thường khoảng -120 đến -160 mV (bên trong âm).
NO₃⁻ là ion mang điện âm.
Theo nguyên lý điện học:
-
Điện tích âm sẽ bị đẩy khỏi vùng âm.
-
Ion âm muốn đi vào môi trường âm là đi “ngược chiều điện thế”.
Chưa kể, nhiều khi nồng độ NO₃⁻ bên trong còn cao hơn bên ngoài.
Như vậy, đưa NO₃⁻ vào trong tế bào là:
-
Ngược gradient nồng độ.
-
Ngược gradient điện thế.
Điều này có nghĩa:
ΔG > 0
→ Không tự phát.
Nếu chỉ dựa vào khuếch tán, quá trình này gần như không thể xảy ra.
3. Chiến lược của thực vật: Không đẩy Nitrat, mà kéo nó đi cùng
Thay vì bơm trực tiếp NO₃⁻ bằng ATP (rất tốn năng lượng), tế bào thực vật dùng một chiến lược tinh tế hơn.
Trước hết, một bơm proton trên màng sinh chất hoạt động liên tục:
H+-ATPase.
Bơm này:
-
Sử dụng ATP.
-
Đẩy H⁺ từ trong ra ngoài.
Hệ quả:
-
Bên ngoài trở nên acid hơn (nồng độ H⁺ cao).
-
Bên trong càng âm hơn (do H⁺ dương bị đẩy ra).
Tế bào đã tạo ra một dạng “pin sinh học” – gọi là điện hóa thế proton.
Đây là nguồn năng lượng dự trữ.
Và protein NRT1.1 không phải là một cái bơm.
Nó là một cánh cổng biết tận dụng dòng proton.
4. Ẩn dụ: Cánh cổng hai nêm xoay quanh trục
Hãy tưởng tượng một cánh cổng không mở như cửa bản lề thông thường.
Nó gồm hai khối hình nêm ghép lại.
Giữa chúng có một trục xoay.
-
Khi mở ra ngoài, mặt ngoài lộ ra.
-
Khi đóng, cả hai mặt bịt kín.
-
Khi mở vào trong, mặt trong lộ ra.
Không bao giờ có chuyện mở cả hai phía cùng lúc.
Đây chính là mô hình:
Alternating Access Model.
Protein không phải là một ống thông suốt.
Nó là một cánh cổng luân phiên.
5. Chu trình vận chuyển – từng bước một
Trạng thái 1: Mở ra ngoài (Outward-open)
Cổng mở về phía đất.
Bên trong túi liên kết còn trống.
H⁺ có xu hướng đi vào vì:
-
Nồng độ ngoài cao.
-
Điện thế trong âm.
Proton là ion nhỏ, di chuyển nhanh và dễ gắn vào vị trí tích điện âm trong protein.
Trạng thái 2: Gắn H⁺
Khi H⁺ gắn vào, cấu hình protein thay đổi nhẹ.
Điện tích nội tại của túi liên kết biến đổi.
Ái lực với NO₃⁻ tăng mạnh.
Đây là bước kích hoạt.
Nếu không có H⁺ gắn trước, NO₃⁻ gần như không thể gắn hiệu quả.
Trạng thái 3: Gắn đủ hai ion
Khi cả H⁺ và NO₃⁻ cùng nằm trong túi:
Protein đạt trạng thái “đầy tải”.
Đây là điều kiện bắt buộc để chuyển cấu hình.
Không đủ hai ion → không xoay.
Trạng thái 4: Đóng kín (Occluded)
Hai nêm xoay vào nhau.
Túi liên kết bị cô lập.
Không còn thông với môi trường ngoài.
Đây là trạng thái trung gian năng lượng cao.
Trạng thái 5: Mở vào trong (Inward-open)
Protein mở về phía tế bào chất.
H⁺ và NO₃⁻ được giải phóng vào bên trong.
Sau đó protein quay về trạng thái ban đầu.
Chu trình lặp lại.
6. Phân tích năng lượng: Ai thực sự kéo ai?
Năng lượng tự do tổng quát của quá trình gồm hai phần:
ΔG_total = ΔG_H + ΔG_NO3
Trong đó:
-
ΔG_H thường âm (thuận lợi).
-
ΔG_NO3 thường dương (bất lợi).
Nếu |ΔG_H| > |ΔG_NO3| → tổng âm
→ vận chuyển xảy ra.
Nói cách khác:
Proton đi xuống dốc năng lượng.
Nó kéo nitrate leo dốc cùng mình.
Giống như một người đi xuống cầu thang, buộc dây kéo một vật nặng lên phía sau.
7. Điều gì quyết định tốc độ vận chuyển?
Có bốn yếu tố chính:
1. pH ngoài
pH thấp → nhiều H⁺ → tăng động lực.
2. pH trong
pH trong cao → chênh lệch lớn → tăng ΔG_H âm.
3. Điện thế màng
Âm hơn → hút H⁺ mạnh hơn.
4. Nồng độ nitrate ngoài
Càng cao → xác suất gắn càng lớn.
Tất cả những yếu tố này phối hợp quyết định tốc độ vận chuyển.
8. Tính hai chiều – một nguyên lý nền tảng
Protein không “biết” chiều đúng.
Nó chỉ tuân theo nhiệt động học.
Nếu:
-
Điện thế yếu.
-
pH không chênh lệch.
-
Nitrate trong quá cao.
Thì:
ΔG_total có thể dương.
Khi đó, hệ thống có thể đảo chiều.
Đây không phải lỗi.
Đó là bản chất vật lý của mọi protein vận chuyển.
9. Vai trò sinh lý: Không chỉ là hấp thu
NRT1.1 không chỉ là transporter.
Nó còn là cảm biến nitrate.
Khi nitrate thấp:
-
Biểu hiện gene tăng.
-
Rễ bên phát triển mạnh.
Khi nitrate cao:
-
Hệ thống điều hòa âm kích hoạt.
Protein này hoạt động như một “transceptor” – vừa vận chuyển vừa cảm nhận tín hiệu.
10. So sánh với hệ ái lực cao
Khi nitrate cực thấp, cây huy động thêm hệ khác:
NRT2.1.
Hệ này:
-
Ái lực cao hơn.
-
Hoạt động mạnh trong điều kiện thiếu N.
-
Cần protein phụ trợ.
Trong khi NRT1.1 có tính lưỡng ái lực (dual-affinity).
11. Ẩn dụ tổng thể: Hệ thống cảng biển và dòng thủy triều
Hãy hình dung:
-
Proton là thủy triều.
-
Nitrate là tàu hàng.
-
Cánh cổng là âu thuyền.
Khi thủy triều lên, nước tràn vào âu thuyền.
Tàu được kéo vào theo.
Không có thủy triều, tàu không thể đi ngược dòng.
Bơm proton tạo thủy triều.
Transporter mở cửa đúng lúc.
Mọi thứ phối hợp chính xác.
12. Ý nghĩa nông nghiệp
Hiểu cơ chế này giúp:
-
Điều chỉnh pH đất hợp lý.
-
Tối ưu phân nitrate.
-
Giảm thất thoát ra môi trường.
-
Tăng hiệu quả sử dụng Nitơ.
Đất quá kiềm → giảm gradient proton.
Đất quá chua → tăng hấp thu nhưng có nguy cơ stress.
Quản lý pH là chìa khóa.
13. Kết luận
Cơ chế đồng vận chuyển H⁺/NO₃⁻ là một ví dụ kinh điển của:
-
Vận chuyển chủ động thứ cấp.
-
Tận dụng năng lượng gradient.
-
Cấu trúc protein biến đổi luân phiên.
Đó không phải là một ống đơn giản.
Đó là một cánh cổng hai nêm xoay quanh trục.
Nó mở – đóng – xoay – giải phóng – lặp lại.
Mỗi chu trình nhỏ bé ấy diễn ra hàng triệu lần trong mỗi tế bào rễ, mỗi giây.
Từ những chuyển động phân tử vô hình đó, toàn bộ cánh rừng được nuôi sống.










Ẩn dụ “chim non há miệng”